




Volume 9, Number 18: 3 May 2006
In a paper recently published in Nature, Reich et al. (2006) describe the first six years' results of a massive FACE study of perennial grassland species conducted at the Cedar Creek Natural History Area in central Minnesota, USA, where 296 field plots planted so as to support the growth of either 1, 4, 9 or 16 grassland species (four C3 grasses, four C4 grasses, four N-fixing legumes and four non-leguminous forbs) on natural ambient-N soil (where N is limiting to growth) or natural soil receiving an extra 4 g N m-2 yr-1 (applied as NH4NO3) have been continually fumigated with either ambient air of approximately 370 ppm CO2 or air enriched to a CO2 concentration of 560 ppm throughout each year's growing season. As a result of what they observed, the nine researchers conclude that "low availability of N progressively suppresses the positive response of plant biomass to elevated CO2." This conclusion, however, is not supported by their data; and several more years of experimentation will likely be required to determine the truth of the matter.
The case for our rebuttal of Reich et al.'s conclusion is extremely simple. As best we can determine from their graph of total plant biomass vs. time in their natural ambient-N soil plots, the percent increase in plant biomass production induced by the extra ~190 ppm of CO2 they supplied to their CO2-enriched plots progressed from 15% in year 1 to 16% in year 2, to 13% in year 3, to 0% in year 4, to 8% in year 5, and to 12% in year 6, as shown in the following figure. Over the first three years of this graph, it is difficult to claim there is any meaningful trend at all in the data. From year 3 to year 4, however, it is patently obvious that the bottom fell out of the aerial fertilization effect, as the percent growth stimulation dropped all the way to zero for some unknown reason. Nevertheless, the CO2-induced growth stimulation reappeared in year 5, rising from 0% to 8%, and it continued its upward climb in year 6, rising from 8% to 12%, which latter value is probably not statistically different from the 13% biomass stimulation recorded in year 3.
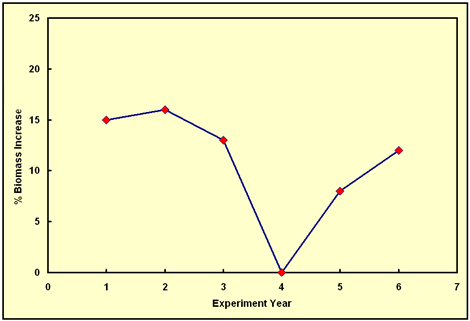
Figure 1. The percent increase in annual biomass production experienced by the plants in the ambient-N soil plots of Reich et al. since the start of their long-term experiment, due to enriching the CO2 concentration of the air above the plots by approximately 190 ppm.
Clearly, the behavior exhibited in the figure above is not that of a low-soil-N-induced "progressive suppression" of the aerial fertilization effect of elevated CO2, because (1) the percent growth enhancement experienced in year 6 of Reich et al.'s study was essentially the same as that experienced in year 3 and (2) the record ends with the aerial fertilization effect in a strong ascending mode. As for years 1 and 2, where the CO2-induced growth stimulation was slightly higher (but only slightly higher), Reich et al. say "the positive response to CO2 enrichment observed under ambient N during the first two years of the experiment may also be more characteristic of seedlings or juvenile plants than of mature well-established plants." This phenomenon (an initial large CO2-induced growth stimulation in the juvenile plant stage that subsequently decreases with time) has been observed in a number of experiments, one of which (Idso and Kimball's long-term sour orange tree study) we have described in some detail in our Editorial of 5 Mar 2003, where it can readily be seen just how difficult - and dangerous - it is to predict the future course of the CO2-induced aerial fertilization effect of atmospheric CO2 enrichment on the basis of even ten years of data.
In light of these several observations, there is no compelling reason to conclude, as Reich et al. do, that "low availability of N progressively suppresses the positive response of plant biomass to elevated CO2." In fact, there is not even a "weak" reason for doing so, as their data depict no such phenomenon. And even if the trend they postulate was apparent, it is clear from the long-term sour orange tree study of Idso and Kimball that six years of data is simply not sufficient to predict the ultimate equilibrium response of a long-lived plant or ecosystem to a large step-increase in atmospheric CO2 concentration in either N-limiting or non-N-limiting situations. On top of that, in the real world of nature the air's CO2 concentration rises gradually, by only a part or two per million per year, which is a much easier situation for plants to deal with than that employed in essentially all CO2 enrichment experiments that have been conducted to date, where a CO2 concentration increase that is fully two orders of magnitude greater than that experienced each year in nature is typically imposed upon the plants.
In conclusion, we encourage Reich et al., as well as those who fund them, to do all they possibly can to keep their experiment going for as long as they possibly can; for the answer they seek is out there, but it is way out there, and their journey to find it has only just begun.
Sherwood, Keith and Craig Idso
Reference
Reich, P.B., Hobbie, S.E., Lee, T., Ellsworth, D.S., West, J.B., Tilman, D., Knops, J.M.H., Naeem, S. and Trost, J. 2006. Nitrogen limitation constrains sustainability of ecosystem response to CO2. Nature 440: 922-925.