




CO2, Global Warming and Coral Reefs
3. Massive, Widespread Coral Bleaching
3.1. Is it Caused by Global Warming?
In an impressive analysis of one of the major environmental questions of our day, Hoegh-Guldberg (1999) concludes that coral bleaching is due to warmer than normal temperatures and that increased sea temperature is the primary reason for why coral bleaching has occurred with increasing intensity and frequency over the past two decades. As outlined in the preceding sections, there is indeed a significant body of evidence that points towards these conclusions; but there is much other evidence that points to alternative possibilities.
Consider, for example, the persistence of coral reefs through geologic time, which provides substantive evidence that these ecological entities can successfully adapt to a dramatically changing global environment (Veron, 1995). What can their history tell us about bleaching and global warming in our day?
The earliest coral reefs date to the Palaeozoic Era, over 450 million years ago (Hill, 1956); while the scleractinian corals, which are the major builders of the reefs of today (Achituv and Dubinsky, 1990), appeared in the mid-Triassic some 240 million years later (Hill, 1956), when the earth was considerably warmer than it is currently (Chadwick-Furman, 1996). Although reef-building ceased for a time following the extinctions at the end of the Triassic, the Scleractinia came back with a vengeance during the Jurassic (Newell, 1971; Veron, 1995); and they continued to exhibit great robustness throughout the Cretaceous, even when temperatures were as much as 8-15°C (Chadwick-Furman, 1996; Veizer et al., 1999), and atmospheric CO2 concentrations 2 to 7 times (Berner and Kothavala, 2001), higher than present.
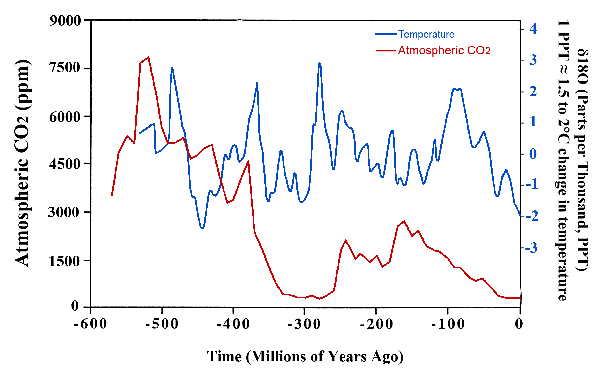
Source: CO2 data from Berner and Kothavala (2001); temperature data from Veizer et al. (1999), updated in 2004 and available at http://www.science.uottawa.ca/~veizer/isotope_data/.
At the end of the Cretaceous, 70% of the genera and one-third of the families of scleractinian corals disappeared (Veron, 1995) in the greatest biospheric extinction event in geological history, which may possibly have been caused by a large asteroid impact (Alvarez et al., 1980, 1984). They developed again, however, throughout the Cenozoic, particularly the Oligocene and Miocene (Chadwick-Furman, 1996). Finally, throughout the past two million years of the Pleistocene, they survived at least seventeen glacial-interglacial cycles of dramatic climate change and sea level fluctuation, successfully adapting, over and over again, to these enormous environmental challenges (Pandolfi, 1999). In the words of Benzie (1999), this evidence suggests that coral reef communities are relatively resilient, have survived previous global climate change, and appear likely to survive future changes. And this conclusion leads us to wonder why corals should be succumbing to global warming now.
To answer such an inquiry we must first address the question of what is normal for coral reefs in our day? Is it what they look like now? Or what they looked like thirty years ago? Or 300 years ago? Or whenever? Kinzie (1999) has emphatically stated that it is clear that the definition of a healthy reef as what it looked like when I started diving is fraught not only with hubris but strong temporal bias. Indeed, as Greenstein et al. (1998a) have observed, it must be demonstrated that the classic reef coral zonation pattern described in the early days of coral reef ecology, and upon which healthy versus unhealthy reefs are determined, are themselves representative of reefs that existed prior to any human influence. Only when this criterion is met will we have, in the words of Greenstein et al. (1998b), a good replacement for the temporally myopic view afforded by monitoring studies that rarely span a scientific career. Clearly, therefore, there should be no argument over the key fact that we need a proper understanding of the past to correctly judge the present if we ever are to foretell the future.
In an attempt to obtain a true picture of pristine coral conditions in the western North Atlantic and Caribbean, Greenstein et al. (1998a, 1998b) conducted systematic censuses of life assemblages and death assemblages of corals on healthy modern patch reefs and compared the results with similar censuses they conducted on fossil assemblages preserved in Pleistocene limestones in close proximity to the modern reefs. What they found was most interesting. The data revealed a recent decline in thickets of Acropora cervicornis, as evidenced by their abundance in the death assemblage, and a concurrent increase in Porites porites, as evidenced by their abundance in the life assemblage. In comparing these results with those obtained from the fossil assemblage, they found that the present Caribbean-wide decline of A. cervicornis is without historical precedent and that it is a dramatic departure from the long-term persistence of this taxon during Pleistocene and Holocene Optimum time, when intensifying cycles in climate and sea level recurred again and again throughout a roughly one-million-year time period.
These observations, along with the similar findings of Jackson (1992) and Aronson and Precht (1997), suggest that if little change in coral community structure occurred throughout the Pleistocene when it was often warmer than it is now (Petit et al., 1999) the recent die-off of A. cervicornis cannot be due to global warming alone, or even primarily; for this particular coral has clearly weathered several major episodes of global warming and elevated water temperatures in the past with no adverse consequences. Neither can the corals die-off be due to the CO2-induced decrease in seawater calcium carbonate saturation state that might possibly be occurring at the present time (see the section following on Coral Calcification); for the airs CO2 content has not risen sufficiently to have caused this parameter to decline enough to significantly impact reef coral calcification rates (Gattuso et al., 1998, 1999), as is also demonstrated by the opportunistic replacement of A. cervicornis by P. porites. In addition, in their detailed reconstruction of the history of calcification rates in massive Porites colonies from Australias Great Barrier Reef, Lough and Barnes (1997) report that the mid-twentieth century had the second highest coral growth rate of the past 237 years. Hence, although A. cervicornis has indeed suffered an extreme decrease in abundance throughout the Caribbean in recent years (Hughes, 1994), its precipitous decline cannot be attributed to either global warming or the direct effects of rising CO2.
In light of these data-driven considerations, Greenstein et al. (1998a, 1998b) have attributed the increasing coral bleaching of the past two decades to a host of local anthropogenic impacts; and this conclusion is accepted in a much wider context as well, as Buddemeier and Smith (1999) have noted that reviews of the problems facing coral reefs have consistently emphasized [our italics] that local and regional anthropogenic impacts are a far greater immediate threat to coral reefs than Greenhouse-enhanced climate change. It is possible, however, that yet another natural phenomenon may be playing an important role on the global stage as well.
3.2. An Alternative Hypothesis
The preceding considerations clearly indicate that global warming cannot be the primary cause of the massive coral bleaching the earth has experienced in recent years. However, many climate alarmists tenaciously cling to this hypothesis because of the fact that (1) no significant massive and widespread coral bleaching was reported in the 1970s and (2) the global warming hypothesis can account for this observation. Specifically, Hoegh-Guldberg (1999) has suggested that the reason why mass bleaching events are not seen prior to 1980 is that increases in sea temperatures have only become critical since in the 1980s, when El Niño disturbances began to exceed the thermal tolerances of corals and their zooxanthellae as a result of global warming increasing the background temperature to which El Niño thermal effects are added.
This reasoning assumes that no other theory is capable of accounting for the fact that modern mass bleaching events did not begin to occur until 1980; and on the basis of this assumption, Hoegh-Guldberg (1999) concludes that the global warming hypothesis must be correct, even in light of the many problems associated with it. This assumption, however, is not true; for there are other ways of satisfying this critical criterion that do account for the lack of bleaching episodes before 1980, which we describe below.
The North Atlantic Oscillation (NAO) is a phenomenon that is responsible for multiannual to decadal variability in Northern Hemispheric climate that is numerically represented by the pressure difference between the Azores high and the Icelandic low (Dugam et al., 1997). It has been documented over the past 350 years in Greenland ice core reconstructions (Appenzeller et al., 1998) and explicitly quantified from 1864 through 1994 via actual pressure records (Hurrell, 1995), which have been updated through 1998 by Uppenbrink (1999).
Plots of these NAO data sets reveal a shift from strong negative index values in the 1950s and 60s to what Hurrell (1995) describes as unprecedented strongly positive NAO index values since 1980. This observation is especially important, for during times of high NAO index values, there is a significant reduction in atmospheric moisture transport across southern Europe, the Mediterranean, and north Africa (Hurrell, 1995); and Richardson et al. (1999) note that this phenomenon has led to the development of prolonged drought in the Sahel region of Africa since the NAO shift to positive index values in 1980.
One consequence of this drought has been a gradual increase in the dust content of the atmosphere, which in some areas has grown to five-fold what was deemed normal prior to this climatic transition (Richardson et al., 1999). Of particular significance to corals is the fact that this airborne dust carries bacteria, viruses and fungi that can kill them; and Pearce (1999) notes that outbreaks of a number of coral diseases have coincided with years when the dust load in the atmosphere was highest. In 1983, for example when the NAO index reached its highest value since 1864 (Hurrell, 1995) and the atmosphere was exceptionally dusty a soil fungus of the Aspergillus genus appeared in the Caribbean, initiating an onslaught of soft coral sea fans that has now destroyed over 90% of them; and Pearce (1999) notes that there are solid scientific reasons for concluding that the speed and pattern of the funguss spread indicates that it could only have arrived on the trade winds from Africa.
In addition to carrying its deadly biological cargo, the positive-NAO-induced airborne dust is rich in iron, which extra supply, in the words of R.T. Barber as quoted by Pearce (1999), "may have spurred the worldwide growth of a variety of invader organisms harmful to coral ecosystems. What is more, such iron-rich dust has the capacity to fertilize algae that compete with zooxanthellae for other scarce nutrients and reef living space. Abram et al. (2003), for example, reported that a massive coral bleaching event that killed close to 100% of the coral and fish in the reef ecosystem of the Mentawai Islands (located southwest of Sumatra, Indonesia, in the equatorial eastern Indian Ocean) in 1997-1998 was brought about by an anomalous influx of iron provided by atmospheric fallout from the 1997 Indonesian wildfires, which they describe as being the worst wildfires in the recorded history of southeast Asia. The enhanced burden of iron, in turn, spawned a large phytoplankton bloom that likely caused the coral and fish death via asphyxiation. In concluding their paper, Abram et al. warn that widespread tropical wildfire is a recent phenomenon, the magnitude and frequency of which are increasing as population rises and terrestrial biomass continues to be disrupted, and by further stating that reefs are likely to become increasingly susceptible to large algal blooms triggered by episodic nutrient enrichment from wildfires, which phenomenon, in their words, may pose a new threat to coastal marine ecosystems that could escalate into the 21st century.
The time-line for the appearance and progression of these several related phenomena matches perfectly with the timeline of the historical buildup of modern coral reef bleaching throughout the 1980s and 90s. This is not to say, however, that these aggregate phenomena comprise the answer to the problem to the exclusion of all other possible causes, even including global warming. We only suggest that they too must be seriously considered in attempts to identify the true cause or causes of this most distressing development in coral reef history.
Thus, although one can make a rational case for coral bleaching being caused by global warming, there are too many pieces of evidence that contradict this hypothesis for it to be deemed the sole, or even primary, cause of this modern curse of reefs. Furthermore, there is at least one alternative explanation and possibly others yet to be described for the observed historical development of massive coral bleaching episodes that are not contradicted by ancillary considerations.