




The progressive nitrogen limitation hypothesis suggests that low concentrations of soil nitrogen will curtail the ability of the productivity-enhancing effect of rising atmospheric CO2 concentrations to maintain increased plant growth and ecosystem carbon sequestration rates over the long term (Hungate et al., 2003; Luo et al., 2004). In this Summary, we explore this possibility as it pertains to earth's grasslands.
Richter et al. (2003) measured gross rates of N mineralization, NH4+ consumption and N immobilization in soils upon which monocultures of Lolium perenne and Trifolium repens had been exposed to ambient (360 ppm) and elevated (600 ppm) concentrations of atmospheric CO2 for seven years in the Swiss FACE study conducted near Zurich. As they describe it, "after seven years of exposure to elevated CO2, gross mineralization, NH4+ consumption and N immobilization in both the L. perenne and the T. repens swards did not show significant differences," and they report that the size of the microbial N pool and immobilization of applied mineral 15N were not significantly affected by elevated CO2. Hence, the five researchers concluded that their observations "did not support the initial hypothesis," which was "that increased below-ground translocation of photoassimilates at elevated pCO2 would lead to an increase in immobilization of N due to an excess supply of energy to the roots and rhizosphere," which would ultimately lead to a reduction in the size of the growth-promoting effect of elevated atmospheric CO2 that is almost always manifest in short-term CO2 enrichment experiments.
In another pertinent study, after five years of exposure of a nutrient-poor low-productivity calcareous grassland to atmospheric CO2 concentrations of 360 and 660 ppm that was conducted in northwestern Switzerland via screen-aided CO2 control or SACC technology (Leadley et al., 1997), Thurig et al. (2003) measured the vegetative and reproductive responses of the various types of plants that comprised the ecosystem. In doing so, they determined that "the effect of elevated CO2 on the number of flowering shoots (+24%, P < 0.01) and seeds (+29%, P = 0.06) at the community level was similar to [the] above ground biomass response." There was also a 42% (P < 0.01) increase in the mean seed number of graminoids and a 33% (P = 0.07) increase in the mean seed number of forbs; and in most species mean seed weight also tended to be greater (+12%, P < 0.01). Consequently, even in what the researchers describe as a nutrient-poor, low-productivity calcareous grassland, atmospheric CO2 enrichment significantly increased both vegetative and reproductive biomass production. In addition, they say it is known from many studies that heavier seeds result in seedlings that "are more robust than seedlings from lighter seeds (Baskin and Baskin, 1998)." Hence, the continued rising of the air's CO2 content would appear to bode well for these nutrient-poor Swiss grasslands, as well as for other low-productivity grasslands around the world.
Three years later, Reich et al. (2006) described the first six years' results of a massive FACE study of perennial grassland species conducted at the Cedar Creek Natural History Area in central Minnesota (USA), where 296 field plots -- planted so as to support the growth of either 1, 4, 9 or 16 grassland species (four C3 grasses, four C4 grasses, four N-fixing legumes and four non-leguminous forbs) on natural ambient-N soil (where N is limiting to growth) or natural soil receiving an extra 4 g N m-2 yr-1 (applied as NH4NO3) -- have been fumigated with either ambient air of ~370 ppm CO2 or air enriched to a CO2 concentration of 560 ppm throughout each year's growing season. As a result of what they observed, the nine researchers concluded that "low availability of N progressively suppresses the positive response of plant biomass to elevated CO2." This conclusion, however, is not supported by their data.
As best we can determine from Reich et al.'s graph of total plant biomass vs. time in their ambient-N plots (our adaptation of which is shown below), the percent increase in plant biomass production induced by the extra ~190 ppm of CO2 they supplied to their CO2-enriched plots progressed from 15% in year 1 to 16% in year 2, to 13% in year 3, to 0% in year 4, to 8% in year 5, and to 12% in year 6. Over the first three years of the study, it is difficult to claim there is any meaningful trend in the data; but in year 4 it is obvious that something drastic happened, as the CO2-induced growth stimulation dropped all the way to zero. Nevertheless, the CO2-induced growth stimulation reappeared in year 5, rising from 0% to 8%; and it continued its upward climb in year 6, rising from 8% to 12%, which latter value is probably not significantly different from the 13% biomass stimulation recorded in year 3.
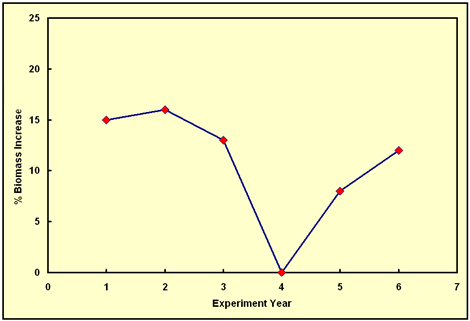
Figure 1. The percent increase in annual biomass production experienced by the plants in the ambient-N soil plots of Reich et al. since the start of their long-term experiment, due to enriching the CO2 concentration of the air above the plots by approximately 190 ppm. Adapted from Reich et al. (2006)
Clearly, the behavior exhibited in the figure above is not that of a low-soil-N-induced "progressive limitation" of the aerial fertilization effect of elevated CO2, because (1) the percent growth enhancement experienced in year 6 of Reich et al.'s study was essentially the same as that experienced in year 3, and (2) the record ends with the aerial fertilization effect in a strong ascending mode. As for years 1 and 2, where the CO2-induced growth stimulation was slightly higher (but only slightly higher), Reich et al. say "the positive response to CO2 enrichment observed under ambient N during the first two years of the experiment may also be more characteristic of seedlings or juvenile plants than of mature well-established plants." This phenomenon (an initial large CO2-induced growth stimulation in the juvenile plant stage that subsequently decreases with time) has been observed in a number of experiments, one of which (Idso and Kimball's long-term sour orange tree study) we have described in some detail in our Editorial of 5 Mar 2003, where it can readily be seen just how difficult -- and dangerous -- it is to predict the future course of the CO2-induced aerial fertilization effect of atmospheric CO2 enrichment on the basis of even ten years of data.
In light of these several observations, there is no compelling reason to believe, as Reich et al. concluded, that "low availability of N progressively suppresses the positive response of plant biomass to elevated CO2." In fact, there is not even a weak reason for doing so, as their data depict no such phenomenon. And even if the trend they postulate was apparent, it is clear from the long-term sour orange tree study of Idso and Kimball that six years of data is simply not sufficient to predict the ultimate equilibrium response of a long-lived plant or ecosystem to a large step-increase in atmospheric CO2 concentration in either N-limiting or non-N-limiting situations. In addition, in the real world of nature the air's CO2 concentration rises gradually, by only a ppm or two per year, which is a much easier situation for plants to deal with than that employed in essentially all CO2 enrichment experiments that have been conducted to date, where a CO2 concentration increase that is fully two orders of magnitude greater than that experienced each year in nature is typically imposed upon the plants being studied all at once. Hence, we would encourage Reich et al., as well as those who fund them, to do all they possibly can to keep their experiment going for as long as they possibly can; for the answer they seek is out there, but it could be way out there, several years in the future.
As another example of the significant amount of time that may be required to adequately evaluate the PNL hypothesis, Zavaleta and Kettley (2006) -- who worked in an annual grassland at the Jasper Ridge Biological Preserve in the interior foothills of the central coast mountain range south of San Francisco, California (USA) -- examined patterns of production, standing biomass, carbon and nitrogen storage, community composition, and soil moisture along a 25-year chronosequence of sites that were in various stages of invasion by the woody shrub Baccharis pilularis. In doing so, they sought evidence for testing the hypothesis that "Baccharis-invaded sites would experience increasing nitrogen limitation as nitrogen was immobilized in biomass and litter," and that this phenomenon would preclude further increases in ecosystem biomass. At the conclusion of their study, however, they learned something quite different.
In progressing from initial grassland conditions to conditions that prevailed 25 years after shrub invasion began, the two researchers found that "net increases in biomass and tissue and soil C:N [carbon to nitrogen ratio] contributed to increases in total ecosystem carbon storage of over 125%." Even more stunning was their discovery that the increases in ecosystem biomass "drove increases in ecosystem nitrogen sequestration of ~700%." In terms of the soil alone, nitrogen content also "increased rapidly with shrub age," since the increase in soil nitrogen "was much larger than the increase in nitrogen immobilization in biomass and litter over time."
What was the source of the extra nitrogen? From whence did it come? The researchers mentioned several possibilities, but could not be sure of them. Nevertheless, their real-world observations clearly repudiate the myopic view, long held by many, that a soil of low initial nitrogen status must constrain long-term positive ecosystem responses to biomass-enhancing phenomena, such as woody plant invasions and atmospheric CO2 enrichment. And in further discussing the subject, Zavaleta and Kettley say that "while many climate models now incorporate the effects of short-term energy and resource exchanges between the atmosphere and the biosphere, most do not consider feedbacks associated with long-term vegetation changes." In this regard, they say their findings "illustrate the potential for important vegetation-mediated ecosystem responses and feedbacks to atmospheric CO2 and climate change," while additionally noting that many of the changes they observed "were progressive and did not saturate with time."
In one final grassland study, Drissner et al. (2007) measured soil microbial biomass and the activities of associated enzymes covering cycles of the most important elements (C, N and P) over a period of nine years in a FACE study that was conducted near Zurich, Switzerland, on three types of grassland (monocultures and mixed cultures of Trifolium repens L. and Lolium perenne L.) that received two different nitrogen treatments (low N = 14 g N m-2 year-1; high N = 56 g N m-2 year-1) and that were maintained at atmospheric CO2 concentrations of either 350 or 600 ppm. This work revealed that that "the enrichment in CO2 increased soil microbial biomass (+48.1%) as well as activities of invertase (+36.2%), xylanase (+22.9%), urease (+23.8%), protease (+40.2%) and alkaline phosphomonoesterase (+54.1%) in spring," and that "in autumn, the stimulation of microbial biomass was 25% less and that of enzymes 3-12% less than in spring," all of which increases, in the researchers' words, "were most likely stimulated by increase in roots under enriched CO2."
Consequently, and in contrast to periodic claims that the future availability of nitrogen will likely be too low to support large CO2-induced increases in plant growth over the long term, primarily because of the contention that when CO2 enrichment increases soil C:N ratios, decomposing microorganisms require more nitrogen, and the claim that this effect can reduce nitrogen mineralization (which is believed to be the main source of nitrogen for plants), Drissner et al. found that in their nine-year-long FACE experiment "stimulation of enzyme activities in the enriched CO2 indicated enhanced C, N and P cycling and greater availabilities of nutrients for microbial and plant growth [our italics]." And they go on to say that their results "support the hypothesis of positive feedback proposed by Zak et al. (1993), who stated that additional C stimulates microbial decomposition and thus leads to more available N under enriched CO2 [our italics]."
Truly, we can expect -- and likely get -- much more from the productivity-enhancing and transpiration-reducing effects of rising atmospheric CO2 concentrations than the world's climate alarmists and biological pessimists have been willing to acknowledge ... and for a much longer period of time.
References
Baskin, C.C. and Baskin, J.M. 1998. Seeds: Ecology, Biogeography, and Evolution of Dormancy and Germination. Academic Press, San Diego, CA.
Drissner, D., Blum, H., Tscherko, D. and Kandeler, E. 2007. Nine years of enriched CO2 changes the function and structural diversity of soil microorganisms in a grassland. European Journal of Soil Science 58: 260-269.
Hungate, B.A., Dukes, J.S., Shaw, M.R., Luo, Y. and Field, C.B. 2003. Nitrogen and climate change. Science 302: 1512-1513.
Leadley, P.W., Niklaus, P.A., Stocker, R. et al. 1997. Screen-aided CO2 control (SACC): a middle ground between FACE and open-top chambers. Acta Oecologica 18: 39-49.
Luo, Y., Su, B., Currie, W.S., Dukes, J.S., Finzi, A., Hartwig, U., Hungate, B., McMurtrie, R.E., Oren, R., Parton, W.J., Pataki, D.E., Shaw, M.R., Zak, D.R. and Field, C.B. 2004. Progressive nitrogen limitation of ecosystem responses to rising atmospheric carbon dioxide. BioScience 54: 731-739.
Reich, P.B., Hobbie, S.E., Lee, T., Ellsworth, D.S., West, J.B., Tilman, D., Knops, J.M.H., Naeem, S. and Trost, J. 2006. Nitrogen limitation constrains sustainability of ecosystem response to CO2. Nature 440: 922-925.
Richter, M., Hartwig, U.A., Frossard, E., Nosberger, J. and Cadisch, G. 2003. Gross fluxes of nitrogen in grassland soil exposed to elevated atmospheric pCO2 for seven years. Soil Biology & Biochemistry 35: 1325-1335.
Thurig, B., Korner, C. and Stocklin, J. 2003. Seed production and seed quality in a calcareous grassland in elevated CO2. Global Change Biology 9: 873-884.
Zak, D.R., Pregitzer, K.S., Curtis, P.S., Teeri, J.A., Fogel, R. and Randlett, D.L. 1993. Elevated atmospheric CO2 and feedback between carbon and nitrogen cycles. Plant and Soil 151: 105-117.
Zavaleta, E.S. and Kettley, L.S. 2006. Ecosystem change along a woody invasion chronosequence in a California grassland. Journal of Arid Environments 66: 290-306.
Last updated 5 November 2008